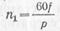Т. к. вода поглощается в осн. корневой системой, возникает необходимость ее передвижения по растению. Этот процесс составляет т. н. восходящий ток. Следует отметить, что это название отражает характер передвижения и его локализацию в растении. Он проходит по мертвым тканям стебля или черешка — сосудам у покрытосеменных и трахеидам у голосеменных. Однако эта локализация не абсолютна: вода способна перемещаться и по флоэмной системе. Передвижение по сосудам идет с большей скоростью (20 см/ч), чем по трахеидам (только 5 см/ч).
Если принять во внимание всю протяженность пути восходящего тока, то его можно будет разбить на 2 не равных по длине участка: Мертвые гистологические элементы в середине проводящего пути — сосуды или трахеиды. Протяженность значительна, но вода проходит по нему сравнительно легко, так как двигается пассивно по мертвым элементам, не испытывая с их стороны значительного сопротивления. Живые клетки корня и листа, находящиеся в начале и в конце пути передвижения. Этот путь пространственно короток, но преодолевается он с большим трудом, так как клеточные мембраны препятствуют передвижению воды. Этот ток снабжает водой все органы и ткани, приводя их в состояние тургора. Восходящий ток воды захватывает мин. ионы, поглощенные корнем, переносит их и тем самым облегчает распределение (но не поглощение!) по растению.
Концевые двигатели водного тока
Для того чтобы вода двигалась по растению (и не просто двигалась, а поднималась вверх), необходимы затраты энергии, точки приложения которой располагаются по концам тока, вследствие чего они получили название концевых двигателей.
Нижний концевой двигатель, или корневое давление. Его роль проявляется в основном при активном поглощении — нагнетании воды. При участии актомиозиноподобных сократительных белков он не только подает воду в корневую систему, но и проталкивает ее дальше в сосуды корня и вверх по стеблю. Нагнетание воды — активный энергозависимый процесс, который сильнее всего выражен в коре корня. Сила, развиваемая концевым двигателем, невелика, она может обеспечить подъем воды на высоту не более 1 м., т. е. достаточна для травянистых растений и небольших кустарников. Верхний концевой двигатель, или присасывающая сила транспирации. При постоянном испарении воды в листьях растений разбивается сосущая сила, отсасывающая воду от ближайших клеток и передающаяся др. кл., по к-ым передвигается вода, вплоть до сосудов. В сосудах нет цитоплазмы, поэтому нет и осмотического давления, и всасывание жидкости происходит при участии всей величины сосущей силы. Она позволяет поднять воду на несколько метров. Этой силы дост. для обеспечения водой кустарников и небольших деревьев.
Поднятие воды по стволу
Многие древесные раст. имеют большую протяженность ствола. У таких растений на помощь приходят силы сцепления между молекулами воды, которые очень велики и могут достигать 30 — 35 МПа. Этой силы достаточно для того, чтобы поднять воду на 1 — 2 км, что значительно превышает высоту любого дерева. Силы сцепления молекул воды действуют только при определенных условиях: водные струи в сосудах должны идти непрерывно, без пузырьков воздуха. Если же в них попадает в-х, передвижение воды прерывается. Непрерывность водных струй в сосудах достигается их сильным натяжением и плотным прилеганием к стенкам сосудов. Водные струи оказываются как бы висящими, они удерживаются в подвешенном состоянии сосущей силой трансп. и снизу поддерживаются силой корневого Р. При сильном натяжении водных струй пр-ит натяжение и самих сосудов, при этом у последних наблюдается увелич. длины и уменьшение поперечного сечения (подобно эластичной резине). Это создает предпосылки к смыканию стенок сосудов, что привело бы к их закрыванию и прекращению водного тока. Однако этого не пр-ит, т. к. сосуды имеют внутренние утолщения стенок — кольчатые, спиральные, лестничные, к-ые наподобие каркаса растягивают их и предохр. от смыкания.
Величина его незначительна — всего 20 см воды на 1 см2 площади сосудов. Однако и этого достаточно при условии не очень сильного напряжения атмосферных факторов.
studfiles.net
Основные двигатели водного тока
Поглощение воды корневой системой идет благодаря работе двух концевых двигателей водного тока: верхнегоконцевого двигателя, или присасывающей силы испарения (транспирации), и нижнего концевого двигателя, или корневого двигателя. Основной силой, вызывающей поступление и передвижение воды в растении, является присасывающая сила транспирации, в результате которого возникает градиент водного потенциала. Водный потенциал – это мера энергии, используемой водой для передвижения. Водный потенциал и сосущая сила одинаковы по абсолютному значению, но противоположны по знаку. Чем меньше насыщенность водой данной системы, тем меньше (более отрицателен) ее водный потенциал. При потере воды растением в процессе транспирации создается ненасыщенность клеток листа водой, как следствие, возникает сосущая сила (водный потенциал падает). поступление воды идет в сторону большей сосущей силы, или меньшего водного потенциала.
Таким образом, верхний концевой двигатель водного тока в растении – это присасывающая сила транспирации листьев, и его работа мало связана с жизнедеятельностью корневой системы. Действительно, опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется.
Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация.
Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, препятствующей испарению, выделяют капельно-жидкую воду с небольшим количеством растворенных веществ – гуттация. Выделение жидкости идет через специальные водные устьица – гидаторы. Выделяющаяся жидкость – гутта. Таким образом, процесс гуттации является результатом одностороннего тока воды, происходящего в отсутствие транспирации, и, следовательно, вызывается какой-то иной причиной.
К такому же выводу можно прийти и при рассмотрении явления плач растений. Если срезать побеги растения и к срезанному концу присоединить стеклянную трубку, то по ней будет подниматься жидкость. Анализ показывает, что это вода с растворенными веществами – пасока. В некоторых случаях, особенно в весенний период, плач наблюдается и при надрезе веток растений. Определения показали, что объем выделяющейся жидкости (пасоки) во много раз превышает объем корневой системы. Таким образом, плач – эт6о не просто вытекание жидкости в результате пореза. Все сказанное приводит к выводу, что плач, как и гуттация, связана с наличием одностороннего тока воды через корневые системы, не зависящего от транспирации. Силу, вызывающую односторонний ток воды по сосудам с растворенными веществами, не зависящую от процесса транспирации, называют корневым давлением. Наличие корневого давления позволяет говорить о нижнем концевом двигателе водного тока. Корневое давление можно измерить, присоединив манометр к концу, оставшемуся после срезания надземных органов растения, или поместив корневую систему в серию растворов различной концентрации и подобрав такую, при которой плач прекращается. Оказалось, что корневое давление равняется примерно 0,1 – 0,15 МПа (Д.А.Сабинин). Определения, проведенные советскими исследователями Л.В.Можаевой, В.Н.Жолкевичем, показали, что концентрация наружного раствора, останавливающего плач, значительно выше концентрации пасоки. Это позволило высказать мнение, что плач может идти против градиента концентрации. Было показано также, что плач осуществляется только в тех условиях, в которых нормально протекают все процессы жизнедеятельности клеток. Не только умерщвление клеток корня, но и снижение интенсивности их жизнедеятельности, в первую очередь интенсивность дыхания, прекращает плач. В отсутствии кислорода, под влиянием дыхательных ядов, при понижении температуры плач приостанавливается. Все сказанное позволило Д.А.Сабинину дать следующее определение: плач растений – это прижизненный односторонний ток воды и питательных веществ, зависящий от аэробной переработки ассимелятов. Д.А.Сабинин предложил схему, объясняющую механизм одностороннего тока воды в корне. Согласно этой гипотезе, клетки корня поляризованы в определенном направлении. Это проявляется в том, что в разных отсеках одной и той же клетки процессы обмена веществ различны. В одной части клетки идут усиленные процессы распада, в частности, крахмала на сахара, вследствие чего концентрация клеточного сока возрастает. На противоположном конце клетки преобладают процессы синтеза, благодаря чему концентрация растворенных веществ в этой части клетки уменьшается. Надо учитывать, что все эти механизмы будут работать только при достаточном количестве воды в среде и не нарушенном обмене веществ.
Согласно другой гипотезе, зависимость плача растений от интенсивности дыхания является косвенной. Энергия дыхания используется для поступления ионов в клетки коры, откуда они десорбируются в сосуды ксилемы. В результате концентрация солей в сосудах ксилемы повышается, что и вызывает поступление воды.
Передвижение воды по растению
Вода, поглощенная клетками корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации, а также силе корневого давления, передвигается до проводящих путей ксилемы. Согласно современным представлениям, вода в корневой системе передвигается не только по живым клеткам. Еще в 1932 г. немецкий физиолог Мюнх развил представление о существовании в корневой системе двух относительно не зависимых друг от друга объемов, по которым передвигается вода, - апопласта и симпласта. Апопласт – это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы. Симпласт – это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему. Апопласт, по-видимому, не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая – по другую сторону клеток эндодермы, и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам Каспари представляют как бы барьер для передвижения воды по свободному пространству (межклетникам и клеточным оболочкам). Для того чтоб попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану и главным образом по апопласту и лишь частично по симпласту. Однако в клетках эндодермы передвижение воды идет, по-видимому, по симпласту. Далее вода поступает в сосуды ксилемы. Затем передвижение воды идет по сосудистой системе корня, стебля и листа.
Из сосудов стебля вода движется через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии. Иначе говоря, в смысле насыщенности водой, имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. Вода передвигается от клетки к клетке благодаря градиенту сосущей силы.
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту значительно большую 10 м. сила сцепления увеличивается, так как молекулы воды обладают большим сродством друг к другу. Силы сцепления обладают и между водой и стенками сосудов.
Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды.
В том случае, если в отдельные членики сосудов попадает воздух, они, по-видимому, выключается из общего тока проведения воды. Таков путь передвижения воды по растению (рис. 1).

Рис. 1. Путь воды в растении.
Скорость перемещения воды по растению в течение суток изменяется. В дневные часы она на много больше. При этом разные виды растений различаются по скорости передвижения воды. Изменение температуры, введение метаболических ингибиторов не влияют на передвижение воды. Вместе с тем этот процесс, как и следовало ожидать, очень сильно зависит от скорости транспирации и от диаметра водопроводящих сосудов. В более широких сосудах вода встречает меньшее сопротивление. Однако надо учитывать, что в более широкие сосуды могут попасть пузырьки воздуха или произойти какие-либо иные нарушения тока воды.
Видео: Движение воды и органических веществ по стеблю. biofile.ru
Т. к. вода поглощается в осн. корневой системой, возникает необходимость ее передвижения по растению. Этот процесс составляет т. н. восходящий ток. Следует отметить, что это название отражает характер передвижения и его локализацию в растении. Он проходит по мертвым тканям стебля или черешка — сосудам у покрытосеменных и трахеидам у голосеменных. Однако эта локализация не абсолютна: вода способна перемещаться и по флоэмной системе. Передвижение по сосудам идет с большей скоростью (20 см/ч), чем по трахеидам (только 5 см/ч).Если принять во внимание всю протяженность пути восходящего тока, то его можно будет разбить на 2 не равных по длине участка: Мертвые гистологические элементы в середине проводящего пути — сосуды или трахеиды. Протяженность значительна, но вода проходит по нему сравнительно легко, так как двигается пассивно по мертвым элементам, не испытывая с их стороны значительного сопротивления. Живые клетки корня и листа, находящиеся в начале и в конце пути передвижения. Этот путь пространственно короток, но преодолевается он с большим трудом, так как клеточные мембраны препятствуют передвижению воды. Этот ток снабжает водой все органы и ткани, приводя их в состояние тургора. Восходящий ток воды захватывает мин. ионы, поглощенные корнем, переносит их и тем самым облегчает распределение (но не поглощение!) по растению.Концевые двигатели водного тока. Для того чтобы вода двигалась по растению (и не просто двигалась, а поднималась вверх), необходимы затраты энергии, точки приложения которой располагаются по концам тока, вследствие чего они получили название концевых двигателей.Нижний концевой двигатель, или корневое давление. Его роль проявляется в основном при активном поглощении — нагнетании воды. При участии актомиозиноподобных сократительных белков он не только подает воду в корневую систему, но и проталкивает ее дальше в сосуды корня и вверх по стеблю. Нагнетание воды — активный энергозависимый процесс, который сильнее всего выражен в коре корня. Сила, развиваемая концевым двигателем, невелика, она может обеспечить подъем воды на высоту не более 1 м., т. е. достаточна для травянистых растений и небольших кустарников. Верхний концевой двигатель, или присасывающая сила транспирации. При постоянном испарении воды в листьях растений разбивается сосущая сила, отсасывающая воду от ближайших клеток и передающаяся др. кл., по к-ым передвигается вода, вплоть до сосудов. В сосудах нет цитоплазмы, поэтому нет и осмотического давления, и всасывание жидкости происходит при участии всей величины сосущей силы. Она позволяет поднять воду на несколько метров. Этой силы дост. для обеспечения водой кустарников и небольших деревьев.Поднятие воды по стволу Многие древесные раст. имеют большую протяженность ствола. У таких растений на помощь приходят силы сцепления между молекулами воды, которые очень велики и могут достигать 30 — 35 МПа. Этой силы достаточно для того, чтобы поднять воду на 1 — 2 км, что значительно превышает высоту любого дерева. Силы сцепления молекул воды действуют только при определенных условиях: водные струи в сосудах должны идти непрерывно, без пузырьков воздуха. Если же в них попадает в-х, передвижение воды прерывается. Непрерывность водных струй в сосудах достигается их сильным натяжением и плотным прилеганием к стенкам сосудов. Водные струи оказываются как бы висящими, они удерживаются в подвешенном состоянии сосущей силой трансп. и снизу поддерживаются силой корневого Р. При сильном натяжении водных струй пр-ит натяжение и самих сосудов, при этом у последних наблюдается увелич. длины и уменьшение поперечного сечения (подобно эластичной резине). Это создает предпосылки к смыканию стенок сосудов, что привело бы к их закрыванию и прекращению водного тока. Однако этого не пр-ит, т. к. сосуды имеют внутренние утолщения стенок — кольчатые, спиральные, лестничные, к-ые наподобие каркаса растягивают их и предохр. от смыкания. Величина его незначительна — всего 20 см воды на 1 см2 площади сосудов. Однако и этого достаточно при условии не очень сильного напряжения атмосферных факторов.
Физиологическая значимость процессов минерального питания, коэффициент накопления, органогены, определение макро- и микроэлементов, их представители. Первичные и вторичные макроэлементы. Тройное правило Арнона.
Растения могут поглощать и перемещать неорганические и органические вещества. Раньше считали, что для роста растений (фотосинтезирующих автотрофов) необходимы только минеральные элементы. Однако в последние годы показано, что многие водоросли требуют витамины В12, а также веществ, содержащих кобальт и т. д.Для наземных растений основным источником воды и минеральных веществ служит почва, не менее важна и атмосфера, которая обеспечивает фотосинтезирующие растения СО2, и, в свою очередь, сама атмосфера поглощает О2 и воду, выделяемую при транспирации. В системе растение-почва-атмосфера происходит значительный круговорот веществ, особенно воды и СО2.Концентрация минеральных веществ в самих растениях в большинстве случаев мало связана с их концентрацией в среде. Например, соотношение K+/Na+ в растениях обычно составляет 5–20, тогда как в среде, как правило, больше Na+. С другой стороны, элементы, содержащиеся в окружающей среде в низких концентрациях, могут накапливаться в растениях в значительном количестве.Эта способность растений характеризуется так называемым коэффициентом накопления:
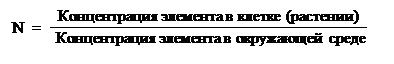 |
Еще раз отметим, что ни присутствие, ни концентрация минеральных элементов в самих растениях не является критерием необходимости. Хорошо зарекомендовало себя в этом отношении (относительно необходимости) так называемое тройное правило Арнона, которое было предложено еще в 1939 году. Элемент признается необходимым в случае, когда: 1. Растение без него не может закончить свой жизненный цикл; 2. Другой элемент не может заменить функцию изучаемого элемента; 3. Элемент непосредственно включен в метаболизм растения.Питательные элементы, которые используются растениями из субстрата в больших количествах (содержание их в золе 10–1–10–2 %), называют макроэлементами, а необходимые в значительно меньшем количестве (содержание 10–3 % и ниже) – микроэлементами.
Вода и СО2 рассматриваются отдельно, но в этом контексте необходимо считать их макроэлементами. К макроэлементам относятся K, Ca, Mg, N, P, S (Н2О и СО2), для галофитов следует добавить Na и Cl. Микроэлементы: Mn, Zn, Cu, Cl, B, Mo, сюда же относится и Fe, который располагается на границе макро- и микроэлементов.Водорослям необходим один или несколько из следующих элементов: Со, Si, I, V. Некоторым высшим растениям необходимы селен, кремний.Кроме необходимых элементов для роста, в оптимальных пределах должны быть и другие факторы: рН, температура, обеспечение О2, градиент водного потенциала, а для зеленых растений – освещенность соответствующей интенсивности и протяженности.Пассивный транспорт – перемещение веществ путем диффузии по градиенту электрохимического потенциала без затраты энергии (простая и, в какой-то мере, облегченная диффузия).Активный транспорт – перемещение веществ против градиента электрохимического потенциала с затратой метаболической энергии, как правило в форме АТФ или редокс-цепей.Для того чтобы понять механизмы трансмембранного переноса элементов минерального питания, остановимся на рассмотрении некоторых физико-химических закономерностях, определяющих движение ионов в растворе и мембране. Начнем с процессов пассивного транспорта (в частности вспомним законы диффузии). Согласно первому закону Фика, поток (Ф) прямо пропорционален коэффициенту диффузии D и градиенту концентрации dC/dх в точке х в данный момент времени.
Ф = – D
cyberpedia.su
Биохимия растений. Процессы брожения. Фотосинтез
теневыносливый растение орошение микрофлора Основным источником влаги является вода, находящаяся в почве, и основным органом поглощения воды в растение-- корневая система. Роль этого органа, прежде всего, заключается в том...
Водний обмін у рослин
Вода відіграє важливу роль у життєдіяльності організму завдяки своїм унікальним фізичним і хімічним властивостям. Молекула води складається з двох атомів водню, приєднаних до одного атому кисню. Атом кисню відтягує електрони від водню...
Водный обмен растений
Поглощение воды корневой системой идет благодаря работе двух концевых двигателей водного тока: верхнего концевого двигателя, или присасывающей силы испарения (транспирации), и нижнего концевого двигателя, или корневого двигателя...
Водный режим растений
Водный режим растений, водообмен, поступление воды в растение и отдача её растением, необходимые для его жизнедеятельности (обмена веществ, роста, развития, размножения)...
Закономерности линейно-весового роста сиговых рыб Обь-Иртышского бассейна
Абсолютная длина тела (общая длина или длина всей рыбы) - от вершины рыла до вертикали конца наиболее длинной лопасти хвостового плавника при горизонтальном положении рыбы. Принимается при указании предельных размеров рыбы...
Кровоносна система
Дослідження ролі вікового чинника у визначенні стійкості риб до отрут важливе для розуміння шляхів впливу токсикантів у природній водоймі на популяцію виду в цілому та на окремі стадії життєвого циклу...
Микоплазмы: представители, особенности строения и метаболизма, систематика
Представители группы микоплазм (роды Mycoplasma, Acholeplasma и Spiroplasma) - паразитические бактерии. Возбудитель относится к разделу Tenericutes. Согласно современной классификации, они относятся к семейству Мycoplasmataceae. Это семейство разделяют на 2 рода - род Мycoplasma...
Морфология внутреннего строения рыб
Основная масса промысловых рыб относится к высшим костным рыбам. Подкласс лучеперых включает более 95 % всех видов современных рыб (треска, окунь, сельдь и др.)- Скелет костных рыб имеет костную ткань...
Особенности строения клетки
В основе расходования воды растительным организмом лежит процесс испарения - переход воды из жидкого в парообразное состояние, происходящий при соприкосновении органов растения с ненасыщенной водой атмосферой...
Пристосування тварин до життя у водному середовищі
Водне середовище має свої певні властивості і тому організми-гідробіонти мають різноманітні специфічні пристосування до життя у воді. Водне середовище менш мінливе, має відносно постійні показники...
Процесс транспирации у разных сортов рода Cucurbita
Вода играет огромную роль в жизни растения. Прямо или косвенно она участвует во всех жизненных процессах, протекающих в клетке. Вода является элементом структуры мембран и цитоплазмы...
Свободные аминокислоты нервной системы
Лизин пока мало исследован в аспекте его значения для нервной системы. Пути деградации лизина в мозге точно не установлены, но они отличаются от локализованных в печени. Лизин в мозге может катаболировать через образование пипеколовой кислоты...
Ткани и их функции в растительных организмах
...
Токсикологія риб
Дослідження ролі вікового чинника у визначенні стійкості риб до отрут важливе для розуміння шляхів впливу токсикантів у природній водоймі на популяцію виду в цілому та на окремі стадії життєвого циклу...
Фитобентос континентальных водоемов
Водяная сеточка (Hydrodictyon reticulatum) Род пресноводных зелёных водорослей порядка протококковых. В СССР -- один вид, Н. reticulatum. Слоевище до 1 м длиной, имеет вид свободноплавающей мешковидной сетки. Рис...
bio.bobrodobro.ru
При низкотемпературных испытаниях можно использовать установку, схематически изображенную на рис. 1.32 [33]. Образец 2 нижним концом закреплен в захвате /. Крутящий момент от двигателя 7 передается образцу с помощью вала 4 через кривошипно-шатунную пару 8 и жесткую пластину 10. Образец вместе с валом 4 совершает знакопеременное кручение. Число циклов регистрируется счетчиком 9. Образец помещают в герметичный криостат 3, снабженный уплотняющим узлом 12 и термопарой 6. Для предварительного охлаждения системы используют сосуд Дьюара 5. Величина крутящего момента измеряется с помощью тензодатчиков 11. [c.46]
В сосуд 16 заливают жидкий азот, а в сосуде 5 через отверстие циркулирует газообразный азот. Для равномерного охлаждения образца битума его помещают в пробирку 14. Для нагрева азота в сосуде 16 помещен нагреватель. Устройство деформации 10 включает две разной длины концентрические трубки 6 и 7 из теплоизоляционного материала с механическим приспособлением 12 для перемещения внутренней трубки вверх и вниз относительно внешней трубки. На нижних концах трубок жестко укреплены захваты с пазами. Приспособление для перемещения внутренней трубки обеспечивает равномерное уменьшение расстояния между пазами захватов на 3,5+0,2 мм от максимального (40+0,1 мм) за 11 + 1 с. Механизм приводится в движение двигателем 13. Для удобства установки пластин предусмотрен фиксатор 11. Аппарат работает в комплекте с потенциометром КСП-4. [c.91]
Для проверки вертикального положения валов этим методом разбирают все направляющие подшипники насоса и электродвигателя, при этом ротор, агрегата оказывается свободно подвешенным на сегментах подпятника. К валу двигателя крепят через изолированную прокладку крестовину, к которой подвешивают струны из калиброванной проволоки диаметром 0,3 0,5 мм с грузом массой 6-15 кг иа конце, опущенным в емкость с вязким маслом. Струны ориентируют по осям насосной станции следующем образом верхний бьеф (+Х), нижний бьеф (-Х), правый берег (+У) и левый берег (- У). Струны опускают так, чтобы они не касались конструкций и висели свободно. Для удобства измерений струны подвешивают на одинаковом расстоянии от вала. С целью повышения точности измерений струны и вал включают в электрическую цепь с напряжением 6-12 В. Струны соединяют через электролампочку или чувствительный гальванометр с одним полюсом батарейки, а вал - с другим полюсом. В момент, когда нутромер касается струны, электрическая цепь замыкается и загорается электрическая лампа или отклоняется стрелка гальванометра. Пог-. решность измерений достигает 0,01 мм. [c.206]
Одним из факторов, сдерживающих возможности форсирования газового двигателя, является работа свечей зажигания. Как показывает практический опыт, для обеспечения нормальной работы свечи зажигания необходимо, чтобы температура нижнего конца её изо- [c.57]
Описанные выше двигатели с принудительным зажиганием и дизели называют четырехтактными за один оборот коленчатого вала происходят впуск и сжатие, за следующий оборот — расщирение и выпуск. Но существуют и двухтактные двигатели. У них некоторые процессы совмещены, и весь цикл протекает за один оборот коленчатого вала. В конце такта расширения открывается выпускное окно, куда выводятся отработавшие газы, и затем открывается впускное окно или впускной клапан, через которые в цилиндр поступает горючая смесь или воздух (в дизеле). Поршень доходит до нижней мертвой точки (крайнее положение при движении поршня в сторону коленчатого вала) и начинает возвращаться к головке цилиндра, перекрывает впускные и выпускные окна, и смесь или воздух в цилиндре сжимается. Перед подходом поршня к верхней мертвой точке в камеру сгорания подается электрическая искра или с помощью форсунки впрыскивается топливо (в дизелях), и вновь начинается процесс расширения. [c.12]
Переключатель П периодически переключается в нижнее положение К, при этом вибропреобразователь оказывается подключенным к сумме напряжения нормального элемента НЭ (или другого стабилизированного источника напряжения) и падения напряжения на эталонном резисторе входящем в мост компенсации температуры свободного конца. Эти напряжения направлены противоположно, поэтому, если они равны и, следовательно, ток моста (а значит, и реохорда) соответствует заданному, то результирующий сигнал равен нулю. Если же ток моста отклонился от заданного значения в ту или иную сторону, то на вибропреобразователе появляется сигнал того или иного знака н реверсивный двигатель, который нри переключении переключателя П от- [c.32]
Отклонение кривошипа относительно его верхнего и нижнего положений в начале открытия или в конце закрытия клапанов неодинаково для разл чных двигателей и зависит от их быстроходности и конструктивных особенностей машины. [c.325]
Передвижение воды по стеблю — ксилемный, или дальний, транспорт — большей частью представляют себе как пассивное движение по непрерывному акропетальному градиенту водного потенциала при участии двух концевых двигателей — нижнего (корневое давление) и верхнего (присасывающее действие транспирации), полагая, что никаких промежуточных двигателей в стебле нет. Правда, движению воды способствует непрерывность водной фазы в растении — от эпидермальных клеток корня до мезофилла листа — и колоссальное натяжение водных нитей в сосудах за счет свойственных воде огромных сил сцепления (см. главу I). Именно благодаря натяжению водных нитей в сосудах и непрерывности водной фазы всякое воздействие на лист, изменяющее скорость транспирации, или на корневую систему, изменяющее скорость поступления воды, влекут за собой мгновенную гидродинамическую реакцию, улавливаемую с помощью чувствительного датчика и аналогичную пульсовой волне в системе кровообращения. Скорость волны во много раз превышает скорость передвижения самой жидкости. Гидродинамические реакции возникают раньше биоэлектрических и, возможно, выполняют в растении даже какую-то информационную роль [337]. Но гидродинамические реакции к промежуточным двигателям непосредственного отношения не имеют. Теория промежуточных двигателей, как отмечает Н. А. Максимов [234], была опровергнута рядом опытов. Так, Е. Ф. Вотчал в своем обширном труде, опубликованном в 1897 г., установил, что вода движется по положенному горизонтально живому отрезку древесного ствола в несколько метров длиной с одинаковой скоростью как от нижнего конца к верхнему, так и наоборот, от верхнего к нижнему и что, следовательно, в древесине отсутствуют какие-либо клапаны, не пропускающие воду вниз а без таких клапанов не могли бы работать предполагаемые двигатели. Подобные же наблюдения были сделаны и другими учеными. Страс-бургеру (1893) и другим исследователям удалось показать, что введенные в перерезанные сосуды ядовитые растворы, например растворы пикриновой кислоты, беспрепятственно поднимаются по древесному стволу на много метров вверх, вплоть до самых листьев, хотя и отравляют на своем пути живые клетки. Точно так же удалось наблюдать беспрепятственное поднятие воды через участки травянистого стебля, убитые нагреванием, замораживанием или действием ядовитых веществ. Через некоторое время, однако, такие убитые участки стеблей пере- [c.147]
Водяной манометр для замера расхода воздуха работает по принципу измерения перепада давления до и после мерной шайбы. Манометр представляет собой стеклянную трубку диаметром 7 мм и длиной более 1 м, укрепленную в металлическом кожухе. В нижней части кожуха имеется резервуар для воды, в верхней— воздушная полость. Стеклянная трубка одним концом соединена с водяным резервуаром, другим — с верхней воздушной полостью. Нижний конец трубки сообщается с воздушной системой перед диафрагмой, верхний конец — с воздушной системой после диафрагмы. В водяной резервуар манометра заливается вода, подкрашенная метиленовым синим. Манометр имеет шкалу с делениями от О до 1000 мм через 2 мм, по которой замеряют уровень воды, соответствующий перепаду давления до и после мерной шайбы. Зная перепад давления, можно по формуле рассчитать количество воздуха, расходуемого двигателем. [c.70]
Направление отклонения конца вала двигателя в плоскости Я определяют по знаку, полученному при подсчете величин 63 и б2у. Если со знаком минус будет 62 , то конец вала имеет отклонение в сторону нижнего бьефа, а если со знаком минус будет 63 , то конец вала отклонен в сторону левого берега. Уклон вала в общем случае может быть расположен между осями X и У, что видно из рис. 14.13, в, и его абсолютное значение получают из соотношения [c.209]
В независимом напорном механизме с отдельным двигателем (рис. 14) возвратно-поступательное движение рукоять получает от двигателя, смонтированного на стреле /. От электродвигателя 2 через две пары цилиндрических зубчатых колес получает вращение вал 5, на концах которого насажены шестерни 4, находящиеся в зацеплении с зубьями реек 5, которые, в свою очередь, закреплены на нижней стороне рукояти ковша. [c.19]
Электродвигатель постоянного тока получает питание от нижнего троллея, проложенного в боковом скрытом туннеле. Пуск электродвигателя и регулирование числа его оборотов осуществляются с площадки управления рубильником 6 и контроллером 7, укрепленным на стойке 8 в конце платформы. Электрическое сопротивление 9, вводимое контроллером в цепь питания, расположено под настилом площадки управления. При подходе к поперечным путям туннельных сушилок платформа затормаживается колодочным тормозом. Последний при включении электродвигателя автоматически растормаживается электромагнитом, а при выключении двигателя затягивается грузом, воздействующим на рычажную систему тормозных колодок. [c.405]
Частота колебаний регулировалась поставленным на приводе вибратора двигателем постоянного тока. Для измерения параметров колебаний использовалась тензометрическая аппаратура и осциллограф или стробоскоп и микроскоп или, в случае колебаний с очень большой амплитудой, стробоскоп и линейка. В нижней части трубы на некотором расстоянии от ее конца помещался нагревательный элемент—медная втулка со-78 [c.78]
Коленчатый вал. Двигатель имеет два коленчатых вала — верхний и нижний. Коленчатые валы отлиты из специального чугуна и термически обработаны. Шейки коленчатых валов пустотелые. Для подвода масла от коренных шеек вала к шатунным в вале сделаны наклонные каналы, в которые вставлены и развальцованы у концов стальные трубки. [c.186]
Рабочий процесс четырехтактного дизеля. Предположим, что поршень 3 двигателя находится в верхнем мертвом положении и при вращении вала 5 начнет двигаться вниз (рис. 125). При этом в цилиндре 2 создается разрежение, газораспределительный механизм открывает впускной клапан 6, и цилиндр заполняется воздухом. Этот такт называется всасыванием (рис. 125, а). К моменту достижения поршнем нижнего крайнего положения прекратится всасывание воздуха и газораспределительный механизм закроет впускной клапан. При движении поршня вверх клапаны впускной 6 и выхлопной 1 закрыты, происходит сжатие воздуха в цилиндре. Этот такт называется тактом сжатия (рис. 125, б). В конце хода сжатия, когда давление воздуха достигает 40 ата, температура его повышается до 600°, через форсунку 7 впрыскивается мелкораспыленное топливо. Попадая в среду сильно разогретого воздуха, топливо быстро воспламеняется и сгорает (рис. 125, в). При горении значительно повышается давление и температура газов. Под давлением этих газов поршень опускается вниз и через шатун 4 передает свое движение коленчатому валу 5. Этот такт называется рабочим ходом. При обратном ходе поршня газораспределительный механизм откроет выхлопной клапан 1, и поршень вытолкнет из цилиндра продукты горения топлива. Этот такт называется выхлопом (рис. 125, г). Далее такты рабочего цикла повторяются. [c.241]
При температуре концов регенераторов минус 70° содержание влаги в воздухе ничтожно мало. Медленно открывая главную задвижку на коллекторе сжатого воздуха, воздух низкого давления подают в регенераторы. Главную задвижку открывают до тех пор, пока давление в нижней колонне поднимется до 4,6—4,8 ати. Если давление продолжает расти, задвижку прикрывают. Одновременно включают все дополнительные сопла турбодетандера, причем сила тока двигатель-генератора не должна превышать 95 а. [c.114]
Общее устройство компрессора показано на рис. 44. На чугунном корпусе 3 расположены цилиндр И и подшипник 6 коренной шейки вала 21. Кривошипная шейка вала 9 соединена с поршнем 10 через кулису. Цилиндр закрыт головкой, состоящей из клапанной плиты 12. по обе стороны которой расположены клапаны — всасывающий 17 и нагнетательный 13. а также крышка головки 14. К нижнему торцу корпуса 3 присоединен статор 20. Ротор 1 с крыльчаткой 2 напрессован на коренную шейку вала. Компрессор с двигателем подвешен в нижней половине кожуха 16 на трех пружинах 4. Пружины прикреплены одним концом к корпусу компрессора 3, а другим — к кронштейнам 5, приваренным к стенкам кожуха. [c.64]
Установлено, что работа этого двигателя на высокосернистом топливе (2,8% серы) и переменных режимах (чередование холостого хода и больших нагрузок) приводит к образованию значительного количества отложений в прорезях маслосъемных колец. При испытании масла без присадки к концу опыта (90 ч) верхнее маслосъемное кольцо забивается на 60%, а нижнее—на 30%. Масла с эффективными присадками значительно снижают интенсивность процесса загрязнения пазов маслосъемных колец. [c.97]
В последние годы получили распространение и электрические турбобуры. Они приводятся в действие специальными электромото- рами малого диаметра, опять-таки помеща- й f емыми в нижнем конце колонны. Энергия подводится к двигателю по специальному кабелю, расположенному внутри бурильной трубы. Такой способ позволяет развивать на долоте значительные усилия, легко поддается автоматизации. [c.47]
На раме ленточного фильтра установлены днище фильтровальной камеры 1 и натяжные ролики 2, несущие бесконечную ленту фильтрующей ткани 13. Над днищем / на прокладке расположена решетка 4, на которую опирается (в рабочей зоне) верхняя ветвь ленты 13, Над решеткой на стойках смонтирована подъемио-опуск-ная крышка 12, имеющая патрубок 6 для ввода суспензии фталевой кислоты и 1,4-нафтохинона. Крышка 12 укреплена на нижнем конце штока 8, верхний конец которого соединен вращающейся гайкой 9 при помощи шлицев с редуктором 15 двигателя 16. При работе двигателя в зависимости от направления вращения гайки 9 крышка 12 поднимается или опускается. Под крышкой 12 расположен неподвижный выталкиватель пасты 5, который укреплен на [c.151]
На комплексе канализационных сооружений г.Зеленограда несколько лет успешно эксплуатируются уровнеиеры песка, разработанные на месте. Уровнемер представляет собой вертикальную штангу, иа нижнем конце которой закреплен дисковый щуп диаметром 150 нм. Верхний конец штанги входит в контактное устройство с микровыключателем, К контактному устройству прикреплен трос, второй конец которого закреплен на барабане многооборотного исполнительного механизма. Когда щуп поднят, штанга под действием собственного веса упирается в нижний упор контактного устройства. Микровыключатель свободен. При опускании штанги щуп упирается в песок, верхний конец штанги перемещается в контактном устройстве, которое опускается исполнительным механизмом. Срабатывает микровыключатель и останавливает двигатель исполнительного механизма. Уровень песка определяется с помощью датчика положения исполнительного механизма. [c.5]
Этот прибор состоит из цилиндра с электрообогревом, соединенного в своем нижнем конце с соплом. Исследуемый расплав с испытательного пуансона выдавливается через сопло в виде жгута, который попадает между двумя зубчатыми колесами, приводимыми в движение двигателем. Зубачатые колеса соединены с чувствительным силомерным устройством. Усилие, приложенное на этот жгут вплоть до его обрыва, регистрируется самописцем. Наличие простой конструкции прибора позволяет производить его точную юстировку. [c.147]
Топливный фильтр тонкой очистки состоит из корпуса 1, опорной плиты 4, верхней крышки 6 и нижней крышки 14. Все эти детали литые, чугунные. Корпус фильтра в нижней части имеет фасонный фланец с полостью 12, в которую поступает нз системы охлаждения двигателя горячая вода, подогревающая топливо в фильтре. Для подвода топлива в верхней части корпуса имеется отверстие, к которому штуцером 15 крепится поворотный угольник тонливоподводящей трубки. В стенке корпуса, примыкающей к фланцу, имеется вертикальный канал И, по которому очищенное топливо подводится к топлнвоотводящей трубке. Снизу к корпусу крепится чугунная крышка 14, имеющая сливное отверстие, которое закрывается пробкой-/5. Сверху корпус закрыт чугунной крышкой, которая вместе с опорной плитой образует полость для прохода очищенного топлива. Продувочный вентиль 5, установленный в верхней крышке, служит для удаления воздуха из фильтра через сливную трубку 3., 1ежду корпусом и верхней крышкой размещается опорная плита, на которой закреплены четыре фильтрующих элемента 2. Фильтрующие элементы надеваются на стержни 9 квадратного сечения и зажимаются между опорной плитой и нижними крышками, которыми заканчиваются стержни. Стержни за верхние концы притягиваются к опорной плите спиральными пружинами/О, которые в сжатом состоянии удерживаются опорными шайбами 7 и штифтами 8. Необходимая герметичность. между корпусом, опорной плитой, верхней и нижней крышка.ми достигается посредством паронитовых прокладок. Внутренние поверхности перед окраской фосфа-тируются. Окраска преследует цель не только предохранить внутренние полости от коррозии, но и предупредить попадание следов формовочной земли и продуктов коррозии в топливо. [c.82]
Все битумные и парафино-канифольные котлы снабжают лопастными мешалками, которые приводятся в движение электрическими двигателями, расположенными в верхней части котлов. Над котлами проходит тельферная грузовая дорога, по которой компоненты изолирующих композиций подвозятся к котлам. На концах трубопроводов в нижней части всех котлов имеются защитные решетки 1, предохраняющие от попадания механических примесей в насосы и трубопроводы. С помощью вентиля 8 и патрубка 17 изолирующие композиции могут отбираться непосредственно из котлов без перекачки их по трубопроводам на сборочные участки. [c.121]
Каждый из них регистрировал одну из компонент вибрации. В точке 4 устанавливался преобразователь, регистрировавший осевые колебания. Вблизи верхнего конца вала насоса с помощью преобразователя 2 регистрировали переходные вибрационные характеристики системы двигатель-насос" при ее пуске и остановке. Один преобразователь, установленный за нижним подшипником вала двигателя, использовался в качестве опорного, относительно его сигналов отсчитьшались фазы и амплитуды остальных датчиков. Результаты измерений, полученные каждым из датчиков, представлялись в виде векторных диаграмм в полярных координатах "амплитуда-фаза". [c.201]
Для программного нагружения кручением в МГТУ им. Баумана спроектирована и опробована установка, изготовленная в Екатеринбурге заводом института ЦНИИМ. Образец располагается вертикально верхним концом он закреплен в тяге, к которой подводится крутящий момент через редуктор от привода, оснащенного двигателем с переменным числом оборотов. Нижний конец образца присоединен к тяге, которая передает прикладываемый к образцу крутящий момент на колесо с двумя гибкими тросиками, присоединенными к динамометрам. По уровню воспринимаемой динамометром силы можно определить крутящий момент, а значит, и напряжение в образце. [c.462]
Колбы с бензином ставят на асбестовую прокладку внутренним диаметром 30 мм, колбы с топливом для реактивных двигателей, керосином, уайт-спиритом, сольвентом и лигроином — на прокладку внутренним диаметром 50 мм, а с дизельным топливом, топливом для тихоходных дизелей и нефтью — на прокладку с внутренним фасонным отверстием 40/50 мм. Отводную трубку келбы соединяют с верхним концом трубки холодильника при помощи плотно пригнанной пробки так, чтобы отводная трубка входила в трубку холодильника на 25—40 мм и не касалась стенок последней. Соединения на корковых пробках заливают коллодием. Затем ставят верхний кожух на асбестовую прокладку, закрывая колбу. При перегонке бензина температура электрического нагревателя и нижнего кожуха не должна быть выше температуры окружающей среды. [c.301]
На штоке клапана у основания тарелки и под тарелкой клапана образуются сравнительно мягкие и липкие отложения, на стеряше клапана, особенно на верхнем его конце, отлагается лак. На тарелке клапана со стороны камеры сгорания наблюдаются твердые отложения в виде нагара, отлагающегося на поршне двигателя, стенках и головке цилиндров, на выпускном клапане и попадающего в зазоры поршневых колец. На последних, а также на нижней поверхности поршня образуется лак. В картере двигателя накапливаются осадки. [c.93]
Топливный насос высокого давления. На двигателях установлены индивидуальные топливные насосы. Регулирование количества подаваемого топлива производится изменением начала и конца 200 400 800 800 1000Ие,з.лс. нодачи посредством поворота плунжера, имеющего на боковой поверхности две спиральные отсечные кромки верхняя — для регулирования начала подачи топлива и нижняя — для регулирования конца нодачи топлива. [c.140]
Прядильные диски установлены на каждом рабочем месте два верхних и два нижних. Верхние диски расположены перпендикулярно фронту обслуживания машины и приводятся во вращение индивидуальными синхронно-реактивными двигателями типа ДРС. Ось нижних прядильных дисков расположена вдоль фронта машины, а сами диски установлены на противоположных концах одного вала, который приводится во вращение зубчатым ремнем от синхроннореактивного двигателя. [c.237]
Секции цепей соединяются между собой осями с насаженными на концах ролика.ми, которые при движении сетки катятся в направляющих. Пластинчатые цепи навешены на две грузовые звездочки, укрепленные на грузовом валу, приводимом во вращение электродвигателем. Секции сетки состоят из рамки и натянутой на нее сетки рамки имеют ребра, предохрзияющие сетку от выпучивания и прорыва. Междурамочное пространство уплотняется специальным козырьком, который вместе с осью звена образует зазор не более 3 мм. Нижнее направляющее устройство имеет радиальные направляющие шины и порог для задержания оседающего песка и ила. Секции сетки через одну и.меют форму ковшиков, что дает возможность в периоды наличия шуги поднимать ее наверх и сбрасывать в отводящий лоток. Промывное устройство состоит из трубы, на которой укреплены два ряда рассеивающих насадок. Вся нагрузка от массы вращающейся сетки воспринимается рамой привода и далее верхним перекрытием водозабора. С целью предохранения полотна сетки от прорыва при перепадах уровня воды бЪлее 30 см работа вращающихся сеток автоматизируется. При достижении предельно допустимого засорения, т. е. перепада, при котором требуется промывка сетки, замыкается контакт дифманометра и открывается задвижка на трубопроводе промывной воды. В результате появления давления на выходе трубопровода, по которому подается промывная вода, замыкается контакт манометра и включается в работу двигатель вращающейся сетки. Таким образом, вначале включается промывка сетки, а затем сетка приводится во вращение. [c.19]
ГАН-8 — гербицидно-аммиачная навесная машина Урожай . Навешивается на тракторы Т-38, Т-16, ДТ-54, ДТ-54А и МТЗ(всех модификаций, самоходные шасси(ДВСШ-16)и может агрегатироваться со всеми навесными и прицепными плугами, культиваторами, а также кукурузной сеялкой СКГН-6. ГАН-8 предназначен для внесения жидкого аммиака в почву, а также для сплошного и ленточного внесения гербицидов. Машина состоит из следующих основных узлов резервуаров, насоса, вакуумной системы заправки, рас-пыливающего устройства, бака для питьевой воды. Два резервуара общей емкостью 560 л укрепляются по бокам трактора. В нижней части каждого резервуара имеется сливной патрубок. Сбоку сливного патрубка расположен штуцер для присоединения всасывающего шланга. Крючки, укрепленные на конце обоих резервуаров, предназначены для закрепления всасывающего шланга (при работе на тракторах Т-38, ДТ-54А). В верхней части резервуара вварен патрубок, на который надевается воздушный шланг. На переднем днище левого резервуара установлено водомерное стекло. Наверху резервуара расположен фиксатор уровня жидкости с краном вакуумного шланга. На переднем днище правого резервуара укреплен кран заборного шланга. В случае монтировки машин на самоходное шасси резервуары между собой соединяют, при этом жидкость к насосу подается из левого резервуара. Насос у ГАН-8 шестеренчатый, приводится в движение от вала отбора мощности трактора. Насос производительностью 60 л в 1 мин., создает давление до 6 кг на 1 см2. Перед запуском насос необходимо заполнить жидкостью. Заправка резервуаров жидкостью производится при помощи вакуумного устройства, которое закрепляется на выхлопной трубе двигателя. Величина вакуума во время работы не регулируется. Время наполнения баков — 3—5 мин. Универсальная штанга ГАН-8 состоит из [c.231]
chem21.info
Силу, вызывающую в растении односторонний ток воды с растворенными веществами, не зависящую от процесса транспирации, называют корневым давлением. Наличие корневого давления позволяет говорить о нижнем концевом двигателе водного тока. Корневое давление можно измерить, присоединив манометр к концу, оставшемуся после срезания надземных органов растения, или поместив корневую систему в серию растворов различной концентрации и подобрав такую, при которой плач прекращается. Оказалось, что величина корневого давления равна примерно 1—1,5 бара.
Было показано также, что плач осуществляется только в тех условиях, в которых нормально протекают все процессы жизнедеятельности клеток корня. Не только умерщвление клеток корня, но и снижение интенсивности их жизнедеятельности, в первую очередь интенсивности дыхания, прекращает плач. В отсутствие кислорода, под влиянием дыхательных ядов, при пониженной температуре плач приостанавливается. Все сказанное позволило Д. А. Сабинину дать следующее определение: плач растений — это прижизненный односторонний ток воды и питательных веществ, связанный с затратой энергии. Однако механизм этого явления до настоящего времени не получил достаточного объяснения. Существуют две основные точки зрения.
Ряд исследователей (А. Крафтс, Т. Бройер и др.) полагают, что вода передвигается пассивно, осмотическим путем по градиенту водного потенциала. Согласно этой гипотезе низкий (более отрицательный) водный потенциал создается в сосудах ксилемы благодаря поступлению туда из почвы растворимых солей, а также из-за слабого противодавления малоэластичных стенок сосудов. Соли поступают активно, для чего и требуется затрата энергии. Таким образом, поступление воды связано в этом случае с энергетическими затратами и с активной жизнедеятельностью клеток корня не прямо, а косвенно. В этой связи корневое давление можно определить как давление, развивающееся в проводящих элементах ксилемы благодаря метаболической активности корней. Поскольку активное поступление зависит от энергии, выделяемой при дыхании, работа нижнего концевого двигателя — корневого давления зависит от:
Вместе с тем существует и другая точка зрения (В.Н. Жолкевич, Л.В. Можаева, Н.В. Пильщиков), согласно которой корневое давление складывается из двух составляющих:
Работа последней требует непосредственной затраты энергии АТФ. Высказывается предположение, что при этом большая роль принадлежит сократительным актиноподобным белкам, энергозависимое сокращение и расслабление которых вызывает изменения гидростатического давления в клетках. В результате на пути водного тока в направлении сосудов ксилемы создаются локальные градиенты водного потенциала, что и способствует проталкиванию воды в сосуды. Вопрос этот требует дальнейшей экспериментальной разработки. Сила, развиваемая нижним концевым двигателем водного тока, во много раз меньше той, которая обусловливается транспирацией. Это проявляется и в том, что скорость вытекания воды из ксилемы значительно уступает скорости транспирации, поэтому в обычных условиях вода в сосудах находится не под положительным, а под отрицательным давлением. Однако все же нижний концевой двигатель способствует подаче воды. Большинство исследователей считает, что прекращение работы нижнего концевого двигателя является показателем начала страдания растений от недостатка воды (засухи). Особенное значение этот двигатель водного тока приобретает в отсутствие транспирации. Так, это может иметь место ранней весной, когда воздух насыщен водой, и транспирация ослаблена. У некоторых многолетних растений в зимний период сосуды ксилемы заполнены воздухом и весной поступление воды идет за счет работы только нижнего концевого двигателя.
vseobiology.ru
Основные двигатели водного тока Поглощение воды корневой системой идет благодаря работе двух концевых двигателей водного тока: верхнего концевого двигателя, или присасывающей силы испарения (транспирации), и нижнего концевого двигателя, или корневого двигателя. Основной силой, вызывающей поступление и передвижение воды в растении, является присасывающая сила транспирации, в результате которого возникает градиент водного потенциала. Водный потенциал – это мера энергии, используемой водой для передвижения. Водный потенциал и сосущая сила одинаковы по абсолютному значению, но противоположны по знаку. Чем меньше насыщенность водой данной системы, тем меньше (более отрицателен) ее водный потенциал. При потере воды растением в процессе транспирации создается ненасыщенность клеток листа водой, как следствие, возникает сосущая сила (водный потенциал падает). поступление воды идет в сторону большей сосущей силы, или меньшего водного потенциала. Таким образом, верхний концевой двигатель водного тока в растении – это присасывающая сила транспирации листьев, и его работа мало связана с жизнедеятельностью корневой системы. Действительно, опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется. Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация. Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, препятствующей испарению, выделяют капельно-жидкую воду с небольшим количеством растворенных веществ – гуттация. Выделение жидкости идет через специальные водные устьица – гидаторы. Выделяющаяся жидкость – гутта. Таким образом, процесс гуттации является результатом одностороннего тока воды, происходящего в отсутствие транспирации, и, следовательно, вызывается какой-то иной причиной. К такому же выводу можно прийти и при рассмотрении явления плач растений. Если срезать побеги растения и к срезанному концу присоединить стеклянную трубку, то по ней будет подниматься жидкость. Анализ показывает, что это вода с растворенными веществами – пасока. В некоторых случаях, особенно в весенний период, плач наблюдается и при надрезе веток растений. Определения показали, что объем выделяющейся жидкости (пасоки) во много раз превышает объем корневой системы. Таким образом, плач – это не просто вытекание жидкости в результате пореза. Все сказанное приводит к выводу, что плач, как и гуттация, связана с наличием одностороннего тока воды через корневые системы, не зависящего от транспирации. Силу, вызывающую односторонний ток воды по сосудам с растворенными веществами, не зависящую от процесса транспирации, называют корневым давлением. Наличие корневого давления позволяет говорить о нижнем концевом двигателе водного тока. Корневое давление можно измерить, присоединив манометр к концу, оставшемуся после срезания надземных органов растения, или поместив корневую систему в серию растворов различной концентрации и подобрав такую, при которой плач прекращается. Оказалось, что корневое давление равняется примерно 0,1 – 0,15 МПа (Д.А.Сабинин). Определения, проведенные советскими исследователями Л.В.Можаевой, В.Н.Жолкевичем, показали, что концентрация наружного раствора, останавливающего плач, значительно выше концентрации пасоки. Это позволило высказать мнение, что плач может идти против градиента концентрации. Было показано также, что плач осуществляется только в тех условиях, в которых нормально протекают все процессы жизнедеятельности клеток. Не только умерщвление клеток корня, но и снижение интенсивности их жизнедеятельности, в первую очередь интенсивность дыхания, прекращает плач. В отсутствии кислорода, под влиянием дыхательных ядов, при понижении температуры плач приостанавливается. Все сказанное позволило Д.А.Сабинину дать следующее определение: плач растений – это прижизненный односторонний ток воды и питательных веществ, зависящий от аэробной переработки ассимелятов. Д.А.Сабинин предложил схему, объясняющую механизм одностороннего тока воды в корне. Согласно этой гипотезе, клетки корня поляризованы в определенном направлении. Это проявляется в том, что в разных отсеках одной и той же клетки процессы обмена веществ различны. В одной части клетки идут усиленные процессы распада, в частности, крахмала на сахара, вследствие чего концентрация клеточного сока возрастает. На противоположном конце клетки преобладают процессы синтеза, благодаря чему концентрация растворенных веществ в этой части клетки уменьшается. Надо учитывать, что все эти механизмы будут работать только при достаточном количестве воды в среде и не нарушенном обмене веществ. Согласно другой гипотезе, зависимость плача растений от интенсивности дыхания является косвенной. Энергия дыхания используется для поступления ионов в клетки коры, откуда они десорбируются в сосуды ксилемы. В результате концентрация солей в сосудах ксилемы повышается, что и вызывает поступление воды.
Передвижение воды по растению Вода, поглощенная клетками корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации, а также силе корневого давления, передвигается до проводящих путей ксилемы. Согласно современным представлениям, вода в корневой системе передвигается не только по живым клеткам. Еще в 1932 г. немецкий физиолог Мюнх развил представление о существовании в корневой системе двух относительно не зависимых друг от друга объемов, по которым передвигается вода, - апопласта и симпласта. Апопласт – это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы. Симпласт – это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему. Апопласт, по-видимому, не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая – по другую сторону клеток эндодермы, и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам Каспари представляют как бы барьер для передвижения воды по свободному пространству (межклетникам и клеточным оболочкам). Для того чтоб попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану и главным образом по апопласту и лишь частично по симпласту. Однако в клетках эндодермы передвижение воды идет, по-видимому, по симпласту. Далее вода поступает в сосуды ксилемы. Затем передвижение воды идет по сосудистой системе корня, стебля и листа. Из сосудов стебля вода движется через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии. Иначе говоря, в смысле насыщенности водой, имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. Вода передвигается от клетки к клетке благодаря градиенту сосущей силы. Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту значительно большую 10 м. сила сцепления увеличивается, так как молекулы воды обладают большим сродством друг к другу. Силы сцепления обладают и между водой и стенками сосудов. Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды. В том случае, если в отдельные членики сосудов попадает воздух, они, по-видимому, выключается из общего тока проведения воды. Таков путь передвижения воды по растению (рис. 1).  Рис. 1. Путь воды в растении.
Рис. 1. Путь воды в растении.
Скорость перемещения воды по растению в течение суток изменяется. В дневные часы она на много больше. При этом разные виды растений различаются по скорости передвижения воды. Изменение температуры, введение метаболических ингибиторов не влияют на передвижение воды. Вместе с тем этот процесс, как и следовало ожидать, очень сильно зависит от скорости транспирации и от диаметра водопроводящих сосудов. В более широких сосудах вода встречает меньшее сопротивление. Однако надо учитывать, что в более широкие сосуды могут попасть пузырьки воздуха или произойти какие-либо иные нарушения тока воды.
Видео: Движение воды и органических веществ по стеблю.
<iframe src="https://www.youtube.com/embed/OfkoOseoD8E" frameborder="0"></iframe>
maxpark.com