Содержание
Сила мышц и способы ее измерения
Дано определение силы скелетных мышц. Описаны способы определения силы скелетных мышц у животных и человека: прямые и косвенные. Дано описание механических динамометров (кистевого и станового), а также электронного динамометра. Приведены результаты прямого измерения силы трехглавой мышцы голени.
Давайте определим, что представляет собой собственно сила мышц и как ее можно измерить?
Определение
Сила (с точки зрения механики) – это векторная величина, мера механического действия на данное тело других тел.
Сила скелетной мышцы (или сила мышечной тяги) – это сила, которая регистрируется на одном из концов мышцы посредством регистрирующего прибора (то есть количественная мера взаимодействия мышцы и регистрирующего прибора).
Более подробно функционирование опорно-двигательного аппарата человека и биомеханика мышц описаны в книге:
Биомеханика опорно-двигательного аппарата человека
Измерение силы мышц
Измерение силы скелетных мышц животных
Для измерения силы скелетных мышц у животных, например, у лягушки, используют специальный прибор (рис. 1). Мышцу подвещивают к одному плечу рычага, а отягощение (W) — к другому.
1). Мышцу подвещивают к одному плечу рычага, а отягощение (W) — к другому.
Рис.1. Измерение силы и скорости сокращения мышц лягушки
Измерение силы скелетных мышц человека
Сила, развиваемая определенной мышечной группой является информативным показателем силовых способностей человека. Для измерения силы скелетных мышц человека используются механические и электронные динамометры, а метод, посредством которого измеряется сила мышц называется динамометрия (динамо — сила, метрия — измеряю).
Важнейшей деталью механических динамометров является пружина, которая должна работать в области линейной деформации. Это означает, что измеряемая сила прямо пропорциональна удлинению пружины. При измерениях в спорте очень часто применяются кистевые (рис.1, 2) и становые (рис. 3) динамометры.
Рис.1. Кистевой динамометр, который использовался в СССРРис.2 Современный кистевой динамометр
Кистевой динамометр позволяет измерить силу мышц кисти человека. Чаще всего его используют для оценки уровня развития физических качеств и состояния здоровья.
Становой динамометр позволяет измерить силу мышц-разгибателей позвоночника. Он в основном применяется в силовых видах спорта (тяжелой атлетике и пауэрлифтинге).
Рис. 3 Становой длинамометр
Недостатком механических динамометров является оценка одного, чаще всего максимального значения силы. Однако, если необходимо изучить изменение усилия, развиваемого мышечной группой или спортсменом в течение определенного отрезка времени, применяются электронные динамометры (рис. 4). В этом случае датчиком является не пружина, а тензодатчик, а сама методика называется тензодинамометрия. На рис. 5 показано измерение силы мышц задней поверхности бедра при помощи электронного динамометра.
Рис.4. Электронный динамометрРис.5. Использование электронного динамометра для измерения силы мышц задней поверхности бедра
Однако измерение силы посредством динамометров обладает рядом недостатков: Во-первых, измеряется сила не одной мышцы, а целой группы мышц, например, сила мышц кисти, силы мышц задней поверхности бедра.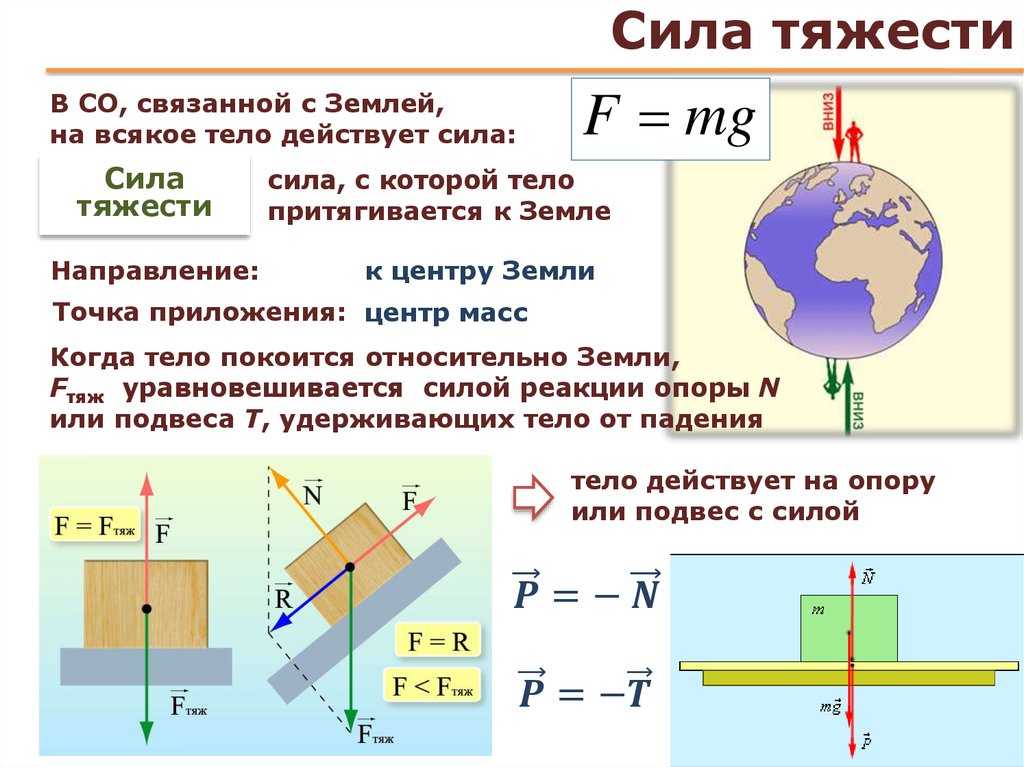 Во-вторых, движения в суставах являются вращательными. Поэтому результат действия мышцы определяет не только сила мышцы, но и плечо силы тяги. То есть на результат, который показывает динамометр еще вляет плечо силы тяги мышцы. Плечо силы тяги мышцы зависит от угла в суставе и места прикрепления мышцы. Поэтому при одном и том же показателе динамометра реальная сила, которую развивает мышца может различаться.
Во-вторых, движения в суставах являются вращательными. Поэтому результат действия мышцы определяет не только сила мышцы, но и плечо силы тяги. То есть на результат, который показывает динамометр еще вляет плечо силы тяги мышцы. Плечо силы тяги мышцы зависит от угла в суставе и места прикрепления мышцы. Поэтому при одном и том же показателе динамометра реальная сила, которую развивает мышца может различаться.
Прямые измерения силы скелетных мышц человека
Ученые давно пытались измерить силу мышц человека при выполнении им двигательных действий. Следует отметить, что напрямую силу мышц у живого человека измерить очень трудно, хотя и возможно. В 2000 году P.V. Komi произвел прямое измерение силы трехглавой мышцы голени при выполнении различных двигательных действий. С этой целью он использовал специальный датчик, который внедряли в ахиллово сухожилие добровольцев (рис. 6). Было установлено, что при беге с максимальной скоростью трехглавая мышца голени может развивать усилие более 8000 Н, а при прыжках — более 3000 Н.
Рис.6 . Демонстрация внедрения датчика в ахиллово сухожилие (P.V.Komi, 2000) для измерения силы трехглавой мышцы
Из-за больших проблем, возникающих при прямом измерении силы мышц при выполнении двигательных действий, для оценки силы, которую развивают мышцы используют косвенные методы, например, моделирование. Также косвенно силу, которую развивает мышца, можно оценить посредством электромиографии.
Литература
- Самсонова А.В. Гипертрофия скелетных мышц человека.– СПб: Кинетика, 2018.– 159 с.
- Бегун П.И., Самсонова А.В. Биомеханика опорно-двигательного аппарата человека: Монография.– СПб: Кинетика, 2020.– 179 с.
- P.V.Komi Stretch-shortening cycle: a powerful model to study normal and fatigued muscle // Journal of Biomechanics, 2000.- Vol.33.- P. 1197-1206.
С уважением, А.В. Самсонова
Похожие записи:
Мышечно-сухожильный комплекс
Приведена рецензия на книгу В.Т.Тураева и В.В. Тюпа «Мышечно-сухожильный комплекс: анатомия, биомеханика, спортивная практика» зав. кафедрой биомеханики НГУ…
кафедрой биомеханики НГУ…
Сила тяжести
Дано определение силы тяжести. Показано, что сила тяжести является частным случаем силы гравитации. Описаны факторы, определяющие силу тяжести:…
Сила
Дано определение силы в механике. Описаны факторы, определяющие действие на тело силы: направление, точка приложения и численное значение.
Типы телосложения (конституции) по Э. Кречмеру
Описана биография Эрнста Кречмера – немецкого психиатра и психолога, разработавшего типологию тела человека. Дано описание типов телосложения…
Типы конституции женщин по И.Б. Галанту
Описана биография известного советского психиатра И.Б.Галанта, предложившего естественную систему конституциональных типов женщин. Дана характеристика предложенных И.Б.Галантом конституциональных…
Звенья тела человека как рычаги
Дано описание опорно-двигательного аппарата (ОДА) человека как системы рычагов. Приведен пример расчета силы двуглавой мышцы плеча…
Зависимость «сила-длина» мышцы
Описаны: история исследования зависимости «сила-длина мышцы», компоненты мышцы, зависимость «сила-длина» расслабленной (пассивной) и активной мышцы; «сила-длина»…
Отсроченное начало болезненности мышц.
 Стратегии лечения и факторы эффективности
Стратегии лечения и факторы эффективности
Описаны симптомы, причины, теории отсроченного начала болезненности мышц (запаздывающих болезненных ощущений, DOMS), а также способы уменьшения этих болей:…
Срочные гормональные ответы у элитных тяжелоатлетов-юниоров
Изучалось изменение концентрации в крови: тестостерона, кортизола, гормона роста, бета-эндорфина и лактата у тяжелоатлетов-юниоров…
Физические качества человека — Департамент физической культуры и спорта
ПечатьDOCPDF
Виктор Николаевич Селуянов, МФТИ, лаборатория «Информационные технологии в спорте»
При движении спортсмена можно зафиксировать его перемещение (положение в пространстве, скорость, ускорение) силу взаимодействия с предметами, и производные переменные — мощность, работа. В педагогике эти физические явления получили иную интерпретацию. Появилось понятие физическое качество и его разновидности — сила, быстрота, выносливость, гибкость и ловкость (В. М. Зациорский, 1966). Для развития этих качеств описываются методы тренировки.
М. Зациорский, 1966). Для развития этих качеств описываются методы тренировки.
Измерить явления, связанные с физической активностью спортсмена возможно, но развивать физические качества нельзя. В организме человека нет физических качеств. В организме есть, например, мышцы, которые могут сокращаться и являть исследователям силу и скорость перемещения костей и тела в целом в пространстве. Для увеличения максимальной силы тяги мышцы требуется изменить строение мышечных волокон (увеличить количество миофибрилл). К сожалению, в педагогической науке все физические явления остаются без глубокого биологического анализа. Спортивная педагогика обособилась, специалисты этой области знаний считают, что у них имеется своя область знаний. Наблюдения в этой области должны составлять основу для образования понятий и положений науки. Это справедливо, в рамках эмпирического изучения объекта исследований. Только надо понимать, что эмпирик признает себя «за полного дурака», ему не доступна суть явлений, он может лишь как пастух в степи петь о том, что видит перед собой. Поэтому в разделе физическая подготовка спортсменов вместо определения смысла физических проявлений занимаются производством новых терминов. Например, способность к прыжкам называют прыгучестью, способность к бегу — бегучесть, способность к ползанию — ползучесть.
Поэтому в разделе физическая подготовка спортсменов вместо определения смысла физических проявлений занимаются производством новых терминов. Например, способность к прыжкам называют прыгучестью, способность к бегу — бегучесть, способность к ползанию — ползучесть.
Давайте рассмотрим обычные педагогические представления о физических качествах с точки зрения биологических наук.
Сила
Под силой понимают способность человека преодолевать внешнее сопротивление за счет активности мышц.
По В. М. Зациорскому (1966) сила человека зависит от:
Педагоги выделяют виды силовых качеств — максимальная сила, скоростная сила, силовая выносливость, взрывную силу, стартовую силу, динамическую силу, статическую силу, эксцентрическая сила. Разумеется, творческие возможности педагогов этим не исчерпываются и можно придумать еще не одну сотню проявления силы, например, в цикле движения во всех видах спорта, что обычно обзывают специальной силой.
С точки зрения биологии и развития силы, в долгосрочной перспективе, максимальное проявление силы зависит от:
— количества мышечных волокон;
— количества миофибрилл в каждом мышечном волокне.
Срочное проявление силы зависит от управления МВ и активности ферментов мышечных волокон.
Центральная нервная система имеет в коре двигательные зоны с нейронами иннервирующими мотонейроны спинного мозга, а те иннервируют мышечные волокна определенную мышцу.
Увеличение силы тяги мышцы связано с рекрутированием двигательных единиц (ДЕ). Каждый двигательный мотонейрон спинного мозга иннервирует множество мышечных волокон, поэтому совокупность «мотонейрон — группа иннервируемых им мышечных волокон», называется двигательная единица.
Каждая двигательная единица имеет свой порог активации (возбуждения) и максимальную частоту. Поэтому при увеличении силы тяги происходит рекрутирование сначала низкопороговых ДЕ, а затем все более высокопороговых ДЕ. При достижении максимальной частоты импульсации мотонейрона мышечные волокна функционируют в режиме тетануса.
У В. М. Зациорского (1966) рассматривается механизм синхронизации работы ДЕ, эту точку зрения сейчас можно рассматривать как ошибку интерпретации физиологических данных. ДЕ работают практически в режиме «все или ничего», иначе говоря, в режиме гладкого тетануса, поэтому нечего синхронизировать. Внутримышечная координация в основном связана с рекрутированием ДЕ с разным порогом активации.
ДЕ работают практически в режиме «все или ничего», иначе говоря, в режиме гладкого тетануса, поэтому нечего синхронизировать. Внутримышечная координация в основном связана с рекрутированием ДЕ с разным порогом активации.
Активность ферментов мышечного волокна зависит от температуры, степени закисления, концентрации адреналина и норадреналина в крови. Этот эффект достигается с помощью разминки (вводной части тренировочного занятия).
Таким образом, срочный механизм управления силой связан с физиологическим законом рекрутирования ДЕ. Способность человека рекрутировать ДЕ существенно различается у мужчин и женщин, молодых и пожилых людей и представителей различных видов спорта. Поддается тренировке с проявлением максимальных силовых возможностей.
Быстрота
Быстроты как физического явления в природе не существует, это обобщающее понятие всех спортивных явлений, которые могут быть описаны как быстрые. Например, различают быстроту простой и сложной двигательной реакции. Эти явления к физике не имеют никакого прямого отношения. А вот скорость сокращения мышцы, темп движений являются физическими явлениями.
Эти явления к физике не имеют никакого прямого отношения. А вот скорость сокращения мышцы, темп движений являются физическими явлениями.
С точки зрения биологии скорость сокращения мышцы зависит от:
— внешнего сопротивления, в соответствии с законом «сила-скорость» Хилла;
— мышечной композиции;
— максимальной силы.
Темп зависит как от скорости одиночного сокращения, так и от скорости расслабления мышц антагонистов. Скорость расслабления зависит от мощности работы кальциевых насосов, а те, в свою очередь, от массы сарколемальных митохондрий.
Выносливость
Под выносливостью понимают способность спортсмена выполнять заданное физическое упражнение без потери мощности, преодолевая утомление.
Педагоги различают общую и специальную выносливость.
Биологи (Н. И. Волков) рассматривают проявление выносливости в зависимости от разных типов энергообеспечения мышечной деятельности и сторон ее проявления:
— алактатная мощность, эффективность и емкость;
— анаэробная гликолитическая мощность, эффективность и емкость;
— аэробная гликолитическая мощность, эффективность и емкость;
— мощность липолиза, эффективность и емкость.
Алактатная мощность зависит от мышечной массы, которая предопределяет запасы АТФ и КрФ, т. е. скоростную и силовую выносливость.
Анаэробная гликолитическая мощность зависит от массы и буферных свойств гликолитических мышечных волокон, окислительных МВ и крови.
Аэробная гликолитическая мощность зависит от массы митохондрий в окислительных и промежуточных мышечных волокнах.
Мощность липолиза зависит от массы митохондрий в окислительных мышечных волокнах.
Надо заметить, что эти представления были прогрессивными в 60–80-е годы, поскольку позволяли внедрять биологическое знание в теорию и практику физического воспитания. В XXI веке эти представления выглядят слишком примитивными. Представлять организм человека в виде пробирки, в которой крутятся шестеренки четырех метаболических процессов некорректно. Модель организма человека (спортсмена) должна быть сложнее. Сейчас она должна, как минимум, включать совокупность мышц пояса верхних и нижних конечностей в каждой мышце надо предусмотреть наличие мышечных волокон разного типа. Сердечнососудистую и дыхательную системы. Блок управления работой этих систем.
Сердечнососудистую и дыхательную системы. Блок управления работой этих систем.
При рассмотрении процессов энергообеспечения на более сложной моделе существенно меняются представления о построении тренировочного процесса. В дальнейшем эти особенности будут рассмотрены более подробно.
Гибкость
Под гибкостью понимают подвижность в суставах. Различают пассивную и активную гибкость, а также анатомическую. Ограничения подвижности могут быть анатомическими, физиологическими и морфологическими.
Анатомические ограничения связаны с упором в кости или мышцы.
Физиологические ограничения связаны с тонусом растягиваемых мышц и рефлексом на растяжение.
Морфологические ограничения связаны с длиной миофибрилл в мышечных волокнах. Миофибриллы имеют разную длину и самые короткие ограничивают подвижность в суставе. Для увеличения подвижности следует разрывать самые короткие миофибриллы.
Новое методическое направление — стретчинг, основано на понимании основных физиологических законов. При растягивании мышцы возникает рефлекс на растяжение. Чем быстрее растягивается активная мышца тем сильнее она сопротивляется благодаря рекрутированию большего числа ДЕ. Поэтому маховые резкие движения приводят к разрывам активных машечных волокон или их миофибрилл. Для снижения травмирующего эффекта в стретчинге предлагают выполнять растяжение легкими рывками с очень маленькой амплитудой. В этом случае рефлекс на растяжение срабатывает, механические нагрузки малы и травм не возникает.
При растягивании мышцы возникает рефлекс на растяжение. Чем быстрее растягивается активная мышца тем сильнее она сопротивляется благодаря рекрутированию большего числа ДЕ. Поэтому маховые резкие движения приводят к разрывам активных машечных волокон или их миофибрилл. Для снижения травмирующего эффекта в стретчинге предлагают выполнять растяжение легкими рывками с очень маленькой амплитудой. В этом случае рефлекс на растяжение срабатывает, механические нагрузки малы и травм не возникает.
Ловкость
Под ловкостью понимают способность человека рационально строить свои двигательные действия в изменяющихся условиях внешней и внутренней среды. В тех случаях, когда внешние условия стабильны, то говорят о координационных способностях.
С точки зрения физики ловкость конечно нельзя рассматривать как физическое качество. Эту проблему следует рассматривать с позиции технической подготовки спортсмена, проблемы формирования двигательных навыков.
Наследуемые аспекты физических качеств
Спортсмены выбирают вид спорта не только по желанию, но и в результате успешности выступления в соревнованиях. Успех в избранном виде спорта во многом определяется наследственной предрасположенностью.
Успех в избранном виде спорта во многом определяется наследственной предрасположенностью.
Проявление силы и темпы ее развития зависят от количества мышечных волокон, мощности функционирования эндокринной системы. Проявление максимальной скорости сокращения мышцы зависит от мышечной композиции. Проявление выносливости связано с активностью ферментов, отвечающих за различные механизмы энергообеспечения. Некоторые из ферментов, например, анаэробного гликолиза (пируватдегидрогеназа, лактатдегидрогеназа и др.) могут плохо функционировать, ограничивать работоспособность в условиях мышечного закисления.
Таким образом, все перечисленные характеристики наследуются и определяют выбор вида спорта в ходе начала спортивной специализации.
Развиваемые аспекты физических качеств
Сила зависит от физиологического поперечника, который преимущественно увеличивается в результате гиперплазии миофибрилл. Скорость сокращения мышцы зависит от роста силы. Темп зависит от массы саркоплазматических митохондрий. Выносливость зависит от массы миофибриллярных митохондрий, а также от количества гликогена и жира в мышечных волокнах.
Выносливость зависит от массы миофибриллярных митохондрий, а также от количества гликогена и жира в мышечных волокнах.
Заключение
Развитие физических качеств не возможно, это бессмысленно. Развитию подлежат структуры клеток организма спортсменов. Для увеличения проявления силы и скорости сокращения мышц нужно заняться гиперплазией миофибрилл, а для увеличения мощности и продолжительности работы следует увеличить гиперплазию митохондрий.
Следовательно, для роста физических возможностей есть два основных пути — рост массы миофибрилл и митохондрий. Остальные факторы также имеют значение, например, масса гликогена и жира в мышечных волокнах, доставка кислорода к мышечным волокнам.
Микроскопия силы тяги для понимания клеточной механотрансдукции
1. Ингбер Д. Механобиология и болезни механотрансдукции. Энн Мед. 2003; 35: 564–577. doi: 10.1080/07853890310016333. [PubMed] [CrossRef] [Google Scholar]
2. Hahn C, Schwartz MA. Механотрансдукция в физиологии сосудов и атерогенезе. Nat Rev Mol Cell Biol. 2009; 10:53–62. doi: 10.1038/nrm2596. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Nat Rev Mol Cell Biol. 2009; 10:53–62. doi: 10.1038/nrm2596. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
3. Choquet D, Felsenfeld DP, Sheetz MP. Жесткость внеклеточного матрикса вызывает усиление интегрин-цитоскелетных связей. Клетка. 1997;88:39–48. doi: 10.1016/S0092-8674(00)81856-5. [PubMed] [CrossRef] [Google Scholar]
4. Фогель В., Шитц М. Восприятие локальной силы и геометрии регулирует функции клеток. Nat Rev Mol Cell Biol. 2006; 7: 265–275. doi: 10.1038/nrm1890. [PubMed] [CrossRef] [Google Scholar]
5. Riveline D, Zamir E, Balaban NQ, et al. Фокальные контакты как механосенсоры: приложенная извне локальная механическая сила вызывает рост фокальных контактов по mDia1-зависимому и ROCK-независимому механизму. Джей Селл Биол. 2001; 153:1175–1186. doi: 10.1083/jcb.153.6.1175. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
6. Yonemura S, Wada Y, Watanabe T, Nagafuchi A, Shibata M. α-Катенин как датчик натяжения, индуцирующий развитие слипчивых соединений. Nat Cell Biol. 2010;12:533–542. дои: 10.1038/ncb2055. [PubMed] [CrossRef] [Google Scholar]
Nat Cell Biol. 2010;12:533–542. дои: 10.1038/ncb2055. [PubMed] [CrossRef] [Google Scholar]
7. Bosveld F, Bonnet I, Guirao B, et al. Механический контроль морфогенеза с помощью пути полярности плоских клеток Fat / Dachsous / Four-joint. Наука. 2012; 336: 724–727. doi: 10.1126/science.1221071. [PubMed] [CrossRef] [Google Scholar]
8. Карри Ф., Адамсон Р. Эндотелиальный гликокаликс: барьер проницаемости и механосенсор. Энн Биомед Инж. 2012;40:828–839. doi: 10.1007/s10439-011-0429-8. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
9. Swift J, Ivanovska IL, Buxboim A, et al. Ядерный ламин-А масштабируется с жесткостью ткани и усиливает дифференцировку, направленную на матрикс. Наука. 2013;341:1240104. doi: 10.1126/science.1240104. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
10. Chen CS, Alonso JL, Ostuni E, Whitesides GM, Ingber DE. Форма клеток обеспечивает глобальный контроль сборки фокальной адгезии. Biochem Biophys Res Commun. 2003; 307: 355–361. дои: 10.1016/S0006-291Х(03)01165-3. [PubMed] [CrossRef] [Google Scholar]
2003; 307: 355–361. дои: 10.1016/S0006-291Х(03)01165-3. [PubMed] [CrossRef] [Google Scholar]
11. Du Roure O, Saez A, Buguin A, et al. Картирование силы при миграции эпителиальных клеток. Proc Natl Acad Sci U S A. 2005;102:2390–2395. doi: 10.1073/pnas.0408482102. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
12. Discher DE, Janmey P, Wang Yl. Тканевые клетки чувствуют и реагируют на жесткость своего субстрата. Наука. 2005; 310:1139–1143. doi: 10.1126/science.1116995. [PubMed] [CrossRef] [Академия Google]
13. Энглер А.Дж., Сен С., Суини Х.Л., Дишер Д.Э. Эластичность матрикса определяет спецификацию линии стволовых клеток. Клетка. 2006; 126: 677–689. doi: 10.1016/j.cell.2006.06.044. [PubMed] [CrossRef] [Google Scholar]
14. Major LG, Holle AW, Young JL, et al. Адаптация к объему контролирует механотрансдукцию стволовых клеток. Интерфейсы приложений ACS. 2019;11:45520–45530. doi: 10.1021/acsami.9b19770. [PubMed] [CrossRef] [Google Scholar]
15. Lo CM, Wang HB, Dembo M, Wang Yl. Движение клеток определяется жесткостью субстрата. Биофиз Дж. 2000;79: 144–152. doi: 10.1016/S0006-3495(00)76279-5. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Lo CM, Wang HB, Dembo M, Wang Yl. Движение клеток определяется жесткостью субстрата. Биофиз Дж. 2000;79: 144–152. doi: 10.1016/S0006-3495(00)76279-5. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
16. Морин Т.Р., младший, Гассем-Заде С.А., Ли Дж. Микроскопия силы тяги в быстро движущихся клетках показывает отдельные роли ROCK и MLCK в механике втягивание. Разрешение ячейки опыта. 2014; 326: 280–294. doi: 10.1016/j.yexcr.2014.04.015. [PubMed] [CrossRef] [Google Scholar]
17. Мирошникова Ю.А., Моув Дж.К., Барнс Дж.М., и соавт. Механика ткани способствует зависимой от IDh2 обратной связи HIF1α-тенасцин C для регуляции агрессии глиобластомы. Nat Cell Biol. 2016;18:1336–1345. дои: 10.1038/ncb3429. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
18. Чирок А., Замир Э.А., Филла М.Б., Литтл К.Д., Ронгиш Б.Дж. Динамика макросборки внеклеточного матрикса у ранних эмбрионов позвоночных. Curr Top Dev Biol. 2006; 73: 237–258. doi: 10.1016/S0070-2153(05)73008-8. [PubMed] [CrossRef] [Google Scholar]
[PubMed] [CrossRef] [Google Scholar]
19. Даллас С.Л., Чен К., Сивакумар П. Динамика сборки и реорганизации белков внеклеточного матрикса. Curr Top Dev Biol. 2006; 75:1–24. doi: 10.1016/S0070-2153(06)75001-3. [PubMed] [CrossRef] [Академия Google]
20. Плотников С.В., Пасапера А.М., Сабасс Б., Уотерман С.М. Колебания силы внутри фокальных спаек опосредуют ощущение жесткости ВКМ, чтобы направлять направленную миграцию клеток. Клетка. 2012; 151:1513–1527. doi: 10.1016/j.cell.2012.11.034. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
21. Zaidel-Bar R, Cohen M, Addadi L, Geiger B. Иерархическая сборка комплексов адгезии клеток и матрикса. Биохим Сок Транс. 2004; 32: 416–420. doi: 10.1042/bst0320416. [PubMed] [CrossRef] [Академия Google]
22. Кавальканти-Адам Э.А., Фольберг Т., Микуле А., Кесслер Х., Гейгер Б., Спатц Дж.П. Распространение клеток и динамика фокальной адгезии регулируются расстоянием между интегриновыми лигандами. Биофиз Дж. 2007; 92: 2964–2974. doi: 10.1529/biophysj.106.089730. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
doi: 10.1529/biophysj.106.089730. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
23. Huebsch N, Arany PR, Mao AS, et al. Использование опосредованных тракцией манипуляций с интерфейсом клетки/матрикса для управления судьбой стволовых клеток. Нат Матер. 2010; 9: 518–526. doi: 10.1038/nmat2732. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
24. McBeath R, Pirone DM, Nelson CM, Bhadriraju K, Chen CS. Форма клеток, натяжение цитоскелета и RhoA регулируют детерминацию линии стволовых клеток. Ячейка Дев. 2004; 6: 483–495. doi: 10.1016/S1534-5807(04)00075-9. [PubMed] [CrossRef] [Google Scholar]
25. Engler AJ, Griffin MA, Sen S, Bonnemann CG, Sweeney HL, Discher DE. Myotubes оптимально дифференцируются на субстратах с тканеподобной жесткостью: патологические последствия для мягких или жестких микроокружений. Джей Селл Биол. 2004; 166: 877–887. doi: 10.1083/jcb.200405004. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
26. Wen JH, Vincent LG, Fuhrmann A, et al. Взаимодействие жесткости матрикса и связывания белков при дифференцировке стволовых клеток. Нат Матер. 2014;13:979–987. doi: 10.1038/nmat4051. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Wen JH, Vincent LG, Fuhrmann A, et al. Взаимодействие жесткости матрикса и связывания белков при дифференцировке стволовых клеток. Нат Матер. 2014;13:979–987. doi: 10.1038/nmat4051. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
27. Dembo M, Wang YL. Напряжения на поверхности раздела клетка-субстрат во время передвижения фибробластов. Биофиз Дж. 1999; 76: 2307–2316. doi: 10.1016/S0006-3495(99)77386-8. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
28. Harris AK, Stopak D, Wild P. Тракция фибробластов как механизм морфогенеза коллагена. Природа. 1981;290:249–251. дои: 10.1038/290249a0. [PubMed] [CrossRef] [Google Scholar]
29. Polacheck WJ, Chen CS. Измерение сил, создаваемых ячейкой: руководство по доступным инструментам. Нат Методы. 2016;13:415–423. doi: 10.1038/nmeth.3834. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
30. Balaban NQ, Schwarz US, Riveline D, et al. Сборка силы и фокальной адгезии: тесная взаимосвязь изучена с использованием эластичных субстратов с микроузором. Nat Cell Biol. 2001; 3: 466–472. doi: 10.1038/35074532. [PubMed] [CrossRef] [Академия Google]
Nat Cell Biol. 2001; 3: 466–472. doi: 10.1038/35074532. [PubMed] [CrossRef] [Академия Google]
31. Вининг К.Х., Муни Д.Дж. Механические силы управляют поведением стволовых клеток при развитии и регенерации. Nat Rev Mol Cell Biol. 2017;18:728–742. doi: 10.1038/nrm.2017.108. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
32. Razafiarison T, Holenstein CN, Stauber T, et al. Сборка лигандов, управляемая поверхностной энергией биоматериала, сильно регулирует механочувствительность и судьбу стволовых клеток на очень мягких субстратах. Proc Natl Acad Sci U S A. 2018;115:4631–4636. doi: 10.1073/pnas.1704543115. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
33. Hur SS, Zhao Y, Li YS, Botvinick E, Chien S. Живые клетки оказывают трехмерное тяговое усилие на свой субстрат. Селл Мол Биоэнг. 2009; 2: 425–436. doi: 10.1007/s12195-009-0082-6. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
34. Del Alamo JC, Meili R, Alonso-Latorre B, et al. Пространственно-временной анализ подвижности эукариотических клеток с помощью улучшенной силовой цитометрии. Proc Natl Acad Sci U S A. 2007;104:13343–13348. doi: 10.1073/pnas.0705815104. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Пространственно-временной анализ подвижности эукариотических клеток с помощью улучшенной силовой цитометрии. Proc Natl Acad Sci U S A. 2007;104:13343–13348. doi: 10.1073/pnas.0705815104. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
35. Разафиарисон Т., Сильван У., Мейер Д., Снедекер Дж.Г. Поверхностная самосборка коллагена влияет на раннюю передачу сигналов остеогенными стволовыми клетками. Adv Healthc Mater. 2016;5:1481–1492. doi: 10.1002/adhm.201600128. [PubMed] [CrossRef] [Google Scholar]
36. Tan JL, Tien J, Pirone DM, Grey DS, Bhadriraju K, Chen CS. Клетки, лежащие на ложе из микроигл: подход к изоляции механической силы. Proc Natl Acad Sci U S A. 2003; 100:1484–1489. doi: 10.1073/pnas.0235407100. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
37. Fu J, Wang YK, Yang MT, et al. Механическая регуляция клеточной функции с помощью геометрически модулированных эластомерных субстратов. Нат Методы. 2010;7:733–736. doi: 10.1038/nmeth. 1487. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
1487. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
38. Wang YK, Yu X, Cohen DM, et al. Передача сигналов, индуцированная костным морфогенетическим белком-2, и остеогенез регулируются формой клеток, RhoA/ROCK и натяжением цитоскелета. Стволовые клетки Dev. 2012;21:1176–1186. doi: 10.1089/scd.2011.0293. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
39. Маскаринец С.А., Франк С., Тиррелл Д.А., Равичандран Г. Количественная клеточная сила тяги в трех измерениях. Proc Natl Acad Sci U S A. 2009;106:22108–22113. doi: 10.1073/pnas.0904565106. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
40. Ruder WC, LeDuc PR. Клетки набирают обороты в 3D. Proc Natl Acad Sci U S A. 2012;109:11060–11061. doi: 10.1073/pnas.1208617109. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
41. Legant WR, Choi CK, Miller JS, et al. Многомерная микроскопия силы тяги выявляет внеплоскостные вращательные моменты вокруг фокальных спаек. Proc Natl Acad Sci U S A. 2013;110:881–886. doi: 10.1073/pnas.1207997110. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Proc Natl Acad Sci U S A. 2013;110:881–886. doi: 10.1073/pnas.1207997110. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
42. Legant WR, Miller JS, Blakely BL, Cohen DM, Genin GM, Chen CS. Измерение механических усилий, оказываемых клетками в трехмерных матрицах. Нат Методы. 2010;7:969–971. doi: 10.1038/nmeth.1531. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
43. Steinwachs J, Metzner C, Skodzek K, et al. Трехмерная силовая микроскопия клеток в биополимерных сетях. Нат Методы. 2016;13:171–176. doi: 10.1038/nmeth.3685. [PubMed] [CrossRef] [Академия Google]
44. Hall MS, Alisafaei F, Ban E, et al. Волокнистая нелинейная эластичность обеспечивает положительную механическую обратную связь между клетками и внеклеточным матриксом. Proc Natl Acad Sci U S A. 2016;113:14043–14048. doi: 10.1073/pnas.1613058113. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
45. Wang N, Tolic-Norrelykke IM, Chen J, et al. Предварительное напряжение ячейки. I. Жесткость и предварительное напряжение тесно связаны в прикрепленных сократительных клетках. Am J Physiol Cell Physiol. 2002; 282:C606–616. doi: 10.1152/ajpcell.00269.2001. [PubMed] [CrossRef] [Google Scholar]
I. Жесткость и предварительное напряжение тесно связаны в прикрепленных сократительных клетках. Am J Physiol Cell Physiol. 2002; 282:C606–616. doi: 10.1152/ajpcell.00269.2001. [PubMed] [CrossRef] [Google Scholar]
46. Марутхамуту В., Сабасс Б., Шварц США, Гардель М.Л. Сила тяги Cell-ECM модулирует эндогенное напряжение в межклеточных контактах. Proc Natl Acad Sci U S A. 2011;108:4708–4713. doi: 10.1073/pnas.1011123108. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
47. Hur SS, del Alamo JC, Park JS, et al. Роли слияния клеток и сдвига жидкости в трехмерных внутриклеточных силах в эндотелиальных клетках. Proc Natl Acad Sci U S A. 2012;109: 11110–11115. doi: 10.1073/pnas.1207326109. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
48. Alonso-Latorre B, Del Alamo JC, Meili R, Firtel RA, Lasheras JC. Компонент колебательной сократительной силы полюса доминирует над силами тяги, создаваемыми мигрирующими амебоидными клетками. Селл Мол Биоэнг. 2011;4:603–615. doi: 10.1007/s12195-011-0184-9. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2011;4:603–615. doi: 10.1007/s12195-011-0184-9. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
49. Tseng Q, Duchemin-Pelletier E, Deshiere A, et al. Пространственная организация внеклеточного матрикса регулирует расположение межклеточных соединений. Proc Natl Acad Sci U S A. 2012;109: 1506–1511. doi: 10.1073/pnas.1106377109. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
50. Mertz AF, Che Y, Banerjee S, et al. Межклеточные адгезии на основе кадгерина организуют силы тяги эпителиальных клеток к матриксу. Proc Natl Acad Sci U S A. 2013;110:842–847. doi: 10.1073/pnas.1217279110. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
51. Liu Z, Tan JL, Cohen DM, et al. Механическая тяговая сила регулирует размер межклеточных соединений. Proc Natl Acad Sci U S A. 2010; 107:9944–9949. doi: 10.1073/pnas.0914547107. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
52. Ng MR, Besser A, Brugge JS, Danuser G. Картирование динамики передачи силы в межклеточных соединениях эпителиальных кластеров.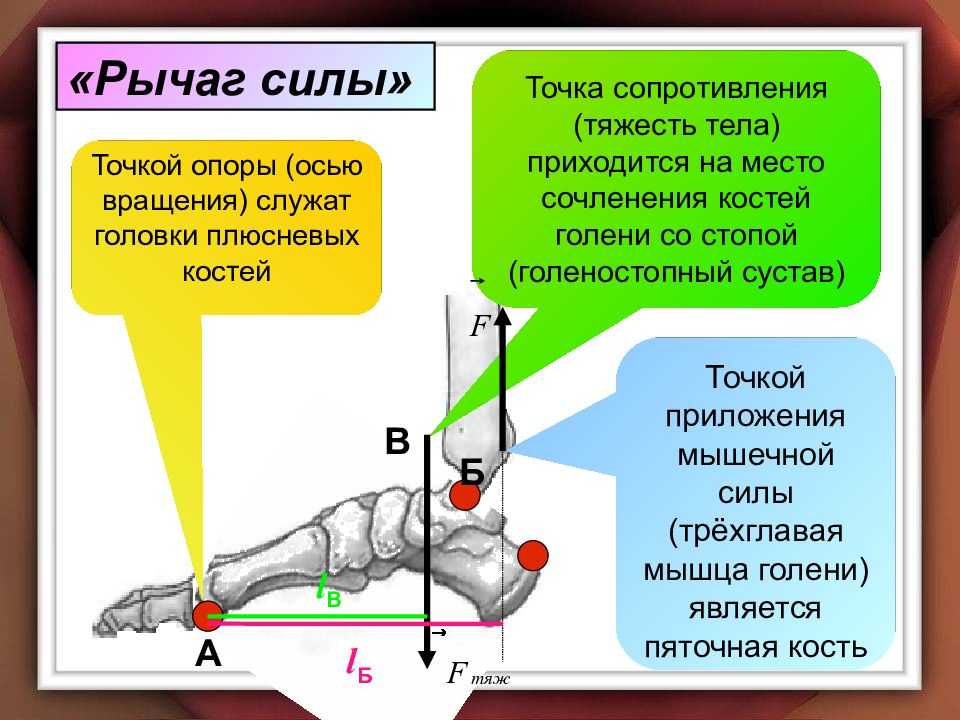 Элиф. 2014;3:e03282. doi: 10.7554/eLife.03282. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Элиф. 2014;3:e03282. doi: 10.7554/eLife.03282. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
53. Tambe DT, Hardin CC, Angelini TE, et al. Коллективное управление клетками совместными межклеточными силами. Нат Матер. 2011; 10: 469–475. doi: 10.1038/nmat3025. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
54. Tambe DT, Croutelle U, Trepat X, et al. Микроскопия монослойного стресса: ограничения, артефакты и точность восстановленных межклеточных напряжений. ПЛОС Один. 2013;8:e55172. doi: 10.1371/journal.pone.0055172. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
55. Serrano R, Aung A, Yeh YT, Varghese S, Lasheras JC, del Alamo JC. Трехмерная монослойная стресс-микроскопия. Биофиз Дж. 2019; 117: 111–128. doi: 10.1016/j.bpj.2019.03.041. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Измерение силы тяги гладкомышечных клеток аорты человека выявило поведение моторного сцепления
Открытый доступ
СТАТЬЯ
Пети Клоди 1 , Гигнандон Ален 2 , Аврил Стефан 1,*
1
Шахты Сент-Этьен, Лионский университет, INSERM, U 1059 SAINBIOSE, F-42023 Сент-Этьен, Франция.
2
Университет Жана Монне, Лионский университет, INSERM, U 1059SAINBIOSE, F-42023 Сент-Этьен, Франция.
* Автор, ответственный за переписку: Аврил Стефан. Электронная почта: .
Молекулярная и клеточная биомеханика 2019 , 16 (2), 87-108. https://doi.org/10.32604/mcb.2019.06415
Резюме
Сократительное поведение гладкомышечных клеток (ГМК) в аорте является важной детерминантой роста, ремоделирования и гомеостаза. Однако количественные значения базального тонуса ГМК никогда точно не характеризовали на отдельных ГМК. Поэтому, чтобы восполнить этот недостаток, мы разработали методику in vitro, основанную на микроскопии силы тяги (TFM). ГМК аорты человека при низких пассажах (4-7) культивировали 2 дня в условиях, способствующих развитию их сократительного аппарата, и высевали на гидрогели с различным модулем упругости (1, 4, 12 и 25 кПа) с внедренными флуоресцентными микросферами. После полной адгезии ГМК искусственно отделяли от геля обработкой трипсином. Движение микрогранул отслеживалось, а поля деформации обрабатывались с помощью механической модели, предполагающей линейную упругость, изотропный материал, плоскую деформацию, для извлечения тяговых сил, ранее прикладываемых отдельными SMC к гелю. Из полученных результатов были сделаны два основных интересных и оригинальных наблюдения о тяговых усилиях SMC: 1. они изменчивы, но обусловлены клеточной динамикой и демонстрируют экспоненциальное распределение с 40–80 % тяговых усилий в диапазоне 0–10 мкН. 2. Они зависят от жесткости подложки: доля сил адгезии ниже 10 мкН имеет тенденцию к уменьшению при увеличении жесткости подложки, тогда как доля более высоких сил адгезии увеличивается. Поскольку эти два аспекта клеточной адгезии (зависимость от изменчивости и жесткости) и распределение их тяговых сил могут быть предсказаны с помощью вероятностной модели двигатель-сцепление, мы делаем вывод, что эта модель может быть применена к ГМК.
После полной адгезии ГМК искусственно отделяли от геля обработкой трипсином. Движение микрогранул отслеживалось, а поля деформации обрабатывались с помощью механической модели, предполагающей линейную упругость, изотропный материал, плоскую деформацию, для извлечения тяговых сил, ранее прикладываемых отдельными SMC к гелю. Из полученных результатов были сделаны два основных интересных и оригинальных наблюдения о тяговых усилиях SMC: 1. они изменчивы, но обусловлены клеточной динамикой и демонстрируют экспоненциальное распределение с 40–80 % тяговых усилий в диапазоне 0–10 мкН. 2. Они зависят от жесткости подложки: доля сил адгезии ниже 10 мкН имеет тенденцию к уменьшению при увеличении жесткости подложки, тогда как доля более высоких сил адгезии увеличивается. Поскольку эти два аспекта клеточной адгезии (зависимость от изменчивости и жесткости) и распределение их тяговых сил могут быть предсказаны с помощью вероятностной модели двигатель-сцепление, мы делаем вывод, что эта модель может быть применена к ГМК.